
蛋白酶体(大分子复合物)
温馨提示:这篇文章已超过415天没有更新,请注意相关的内容是否还可用!
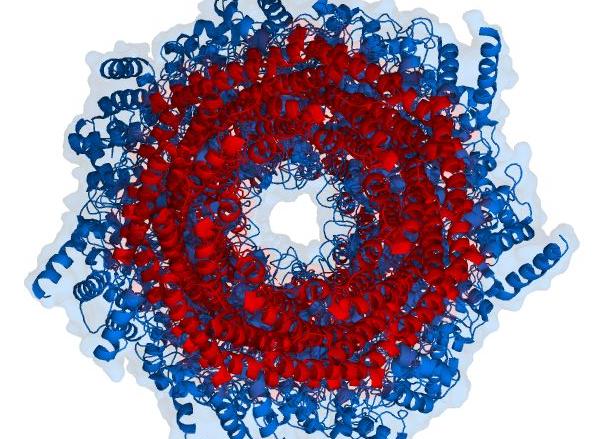
蛋白酶体
大分子复合物
蛋白酶体是一种巨型蛋白质复合物。它既存在于细胞核中,又存在于胞质溶胶中,是溶酶体外的蛋白水解体系,由10~20个不同的亚基组成中空的圆桶形的结构,显示多种肽酶的活性,能够从碱性、酸性和中性氨基酸的羧基侧水解多种与遍在蛋白连接的蛋白质底物。蛋白酶体对蛋白质的降解是与环境隔离的。
主要降解两种类型的蛋白质:一类是错误折叠的蛋白质,另一类就是需要进行数量调控的蛋白质。蛋白酶体对蛋白质的降解通过泛素(ubiquitin)介导,所以又称为泛素降解途径。蛋白酶体对蛋白质的降解作用分为两个过程:一是对被降解的蛋白质进行标记,由泛素完成;二是蛋白酶解作用,由蛋白酶体催化。蛋白酶体存在于所有真核细胞中,其活性受γ干扰素的调节。
| 中文名 | 蛋白酶体 |
| 外文名 | proteasomes |
结构和组成
蛋白酶体结构复杂且具有多种存在形式,其组成可分为核心颗粒(CP)和调节颗粒(RP)两个部分。
核心颗粒
核心颗粒是沉降系数为20S的蛋白酶复合体,又称20S蛋白酶体。它由α环和β环各两个组成,每环各有7个亚基,按α-β-β-α排列成圆桶状结构。其中α环位于两端,β环位于中间,α环中心有一孔道与圆桶内部相连,但α亚基的N末端肽链将此孔道完全封闭,防止不该被降解的蛋白质进入圆桶内部而被水解破坏。
蛋白酶体属于N末端亲核水解酶超级家族,其N末端苏氨酸残基在蛋白水解过程中作为亲核基团进攻肽键,虽然活性位点都是苏氨酸残基,但不同β亚基的催化活性各不相同。α亚基没有催化活性,β环内表面的β1(Y)、β2(Z)和β5(X)亚基具有明显的催化活性,分别为胱天蛋白酶样活性、胰蛋白酶样活性、糜蛋白酶样活性,负责切断酸性、碱性、疏水氨基酸羧基端的肽链。
哺乳动物的蛋白酶体含有7个α亚基和10个β亚基,这是由于在免疫调节细胞因子γ干扰素的诱导作用下,β1、β2、β5亚基可分别为有催化活性的β1i(LMP2)、β2i(MECL-1)、β5i(LMP7)亚基所替代,形成免疫蛋白酶体。这种变化使蛋白酶体降解蛋白质的速度和切割位点与原来相比发生很大变化,发生这种结构变化的机制尚不明确,但其结果更有利于内源性蛋白质的降解和抗原的递呈。
调节颗粒
底物需要通过α环的中心孔道才能进入20S蛋白酶体内部的催化位点,然而在没有调节颗粒存在的情况下孔道是封闭的。调节颗粒结合在20S蛋白酶体的两端,可以使孔道开放且对底物有高度的选择性。蛋白酶体有多种调节颗粒,包括PA700、PA28、PA200等。PA700具有多种生物功能,如开放孔道、识别底物、底物去折叠及泛素的再循环等。
PA700的结构可分为基底部和盖部两部分,基底部位于20S蛋白酶体的两端含有6个三倍体ATP酶亚基(Rpt1-6)和三个非ATP酶亚基(Rpn1、Rpn2、Rpn10),盖部由其他的非ATP酶组成。基底部的ATP酶亚基在ATP参与的情况下与20S蛋白酶体结合,使α环口的孔道打开,而Rpn10则是蛋白酶体上的多聚泛蛋白底物的受体。盖部的作用尚不明确,它可能在多聚泛蛋白从前体到降解的过程中起重要作用。
PA28也可与20S蛋白酶体结合,开放α环口的中心孔道,刺激肽进入和离开蛋白酶体的催化腔。与PA700不同,PA28与20S蛋白酶体的结合不需要ATP的参与。PA28包括3种亚基:PA28α、PA28β和PA28γ,其中PA28α/β在免疫应答过程中起重要作用。有人发现细胞内存在一种杂化蛋白酶体,其核心颗粒两端分别结合一个PA700和一个PA28。
PA200是另一种蛋白酶体的调节颗粒,它与20S蛋白酶体形成复合物,可开放中心孔道加速肽的降解。与PA28相似,PA200与20S蛋白酶体的结合也不需要ATP的参与,另外PA200还在DNA修复过程中起重要作用。
对于20S蛋白酶体的3个催化亚基β1、β2和β5来说,PA700能将每个催化亚基的活性提高3倍;PA28α/β能将3个亚基的活性提高10~20倍,PA28γ能提高β2亚基的活性,而同时却抑制β5亚基的活性;PA200对β1亚基活性的提高是对β2和β5亚基的3倍,这可能是由于PA200导致β1亚基构象发生变化的原因。
降解过程
泛素化和定靶
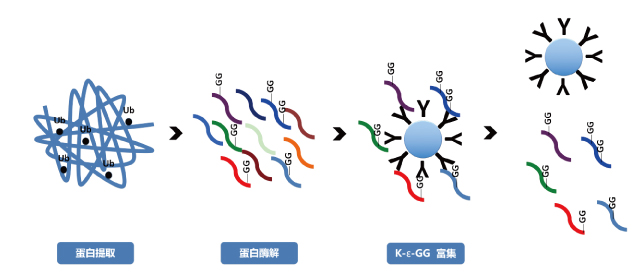 修饰蛋白质组-泛素化
修饰蛋白质组-泛素化需要被蛋白酶体降解的蛋白质会先被连接上泛素作为标记,即蛋白质上的一个赖氨酸与泛素之间形成共价连接。这一过程是一个三酶级联反应,即需要有由三个酶催化的一系列反应的发生,整个过程被称为泛素化信号通路。
在第一步反应中,泛素活化酶(又被称为E1)水解ATP并将一个泛素分子腺苷酸化。接着,泛素被转移到E1的活性中心的半胱氨酸残基上,并伴随着第二个泛素分子的腺苷酸化。被腺苷酸化的泛素分子接着被转移到第二个酶,泛素交联酶(E2)的半胱氨酸残基上。最后,高度保守的泛素连接酶(E3)家族中的一员(根据底物蛋白质的不同而不同)识别特定的需要被泛素化的靶蛋白,并催化泛素分子从E2上转移到靶蛋白上。靶蛋白在被蛋白酶体识别之前,必须被标记上至少四个泛素单体分子(以多泛素链的形式)。
因此,是E3使得这一系统具有了底物特异性。E1、E2和E3蛋白的数量依赖于生物体和细胞类型,人体中就存在大量不同的E3蛋白,这说明泛素-蛋白酶体系统可以作用于数量巨大的靶蛋白。
多泛素化后的蛋白质是如何被蛋白酶体所识别的,还没有完全弄清。泛素受体蛋白的N末端具有一个类泛素结构域,以及一至多个泛素结合结构域。类泛素结构域可以被19S调节颗粒所识别,而泛素结合结构域可以通过形成三螺旋束来结合泛素。这些受体蛋白可能能够结合多泛素化的蛋白质并将其携带到蛋白酶体,而关于这种结合的特异性和调控机制还不清楚。但最近有研究者发现,调节颗粒上的亚基Rpn13可以发挥泛素受体的功能。
泛素蛋白自身由76个残基所组成,以“泛素”为名是因为它在生物体中广泛存在:具有高度保守的序列并且存在于所有已知的真核生物体中。真核生物中编码泛素的基因以串联重复(英语:tandem repeat)的方式排列,这可能是因为大量转录的需要,为细胞生产足够多的泛素。有人提出泛素是目前发现的进化速度最慢的蛋白质。
去折叠和移位
泛素化信号通路。其中,“Ub”表示泛素。
泛素化后的蛋白质(以下称为底物蛋白)被19S调节颗粒所识别,这一过程是一个ATP依赖的结合过程。然后,底物蛋白必须进入20S核心颗粒的内部孔道,以便与位于其中的水解活性位点接触。由于20S颗粒的孔道相对狭窄,而且两端由α环中亚基的N末端控制开关,所以底物蛋白在进入核心颗粒之前必须至少部分去折叠。将去折叠的蛋白质传递进入核心颗粒的过程被称为“移位”(translocation),而移位必须发生在去泛素化之后。但目前对于底物蛋白的去泛素化和去折叠机制还不了解。
在整个降解反应过程中,那一步是限速步取决于底物蛋白的类别;对于一些蛋白质,去折叠过程是限速步,而对于另一些蛋白质,可能是去泛素化为限速因子。至于哪些底物蛋白在移位之前必须去折叠,还未有结论,而牢固的三级结构和一些特殊的非局部相互作用,如二硫键,能够抑制降解。
由α亚基所形成的“门”可以阻止长于四个残基的多肽进入20S颗粒的内部。在识别步骤开始前结合上的ATP分子在移位发生前被水解,而对于水解产生的能量是用于蛋白质去折叠还是“门”的打开还有争议。26S蛋白酶体在存在无法水解的ATP类似物(即无法获得水解产生的能量)的情况下,依然可以降解去折叠的蛋白质,但却无法降解折叠的蛋白质;这一结果说明ATP水解所产生的能量至少部分被用于蛋白质去折叠。在19S帽子处于ATP结合状态时,去折叠的底物蛋白可以由促进扩散作用,传递通过开启的“门”。
球蛋白去折叠的机制是基本类似的,但在一定程度上也取决于蛋白质的氨基酸序列。研究者发现含有较长的甘氨酸或丙氨酸序列可以抑制去折叠,从而降低蛋白酶体的降解效率;其结果是生成含有部分去折叠蛋白质的混合物,这可能是由于ATP水解和去折叠步骤之间的脱节所导致的。
自然界中的一些蛋白质也有这样的甘氨酸-丙氨酸重复序列存在,如蚕丝中的丝心蛋白(fibroin);值得一提的是,特定的人类疱疹病毒基因的表达产物也含有这样的序列,通过抑制蛋白酶体的作用,阻止了抗原呈递到主要组织相容性复合体上,从而有助于病毒的繁殖。
20S核心颗粒的一个剖面图,显示了活性位点的位置。其中,α亚基用绿色的球来表示,β亚基的蛋白骨架显示为飘带,并且不同的多肽链用不同的颜色表示。小的粉色球表示每个亚基的活性位点中苏氨酸残基的位置。淡蓝色的化学结构为结合在活性位点上的抑制剂硼替佐米。
蛋白质的降解
泛素蛋白酶体系统是由泛素介导的一种高度复杂的蛋白降解机制,它参与降解细胞内许多蛋白质并且这个过程具有高度特异性。细胞内蛋白质的降解参与调节许多细胞过程,包括细胞周期、DNA修复、细胞生长和分化、细胞质量的控制、病原生物的感染反应和细胞凋亡等。
蛋白质的降解由20S核心颗粒中的β亚基进行,其机制被认为是苏氨酸依赖的亲核攻击。这一机制可能需要有一个结合的水分子参与活性的苏氨酸上羟基的去质子化。降解发生在核心颗粒中间的两个β环内的孔道里,一般不生成部分降解的产物,而是将底物蛋白完全降解为长度一定的肽段;肽段的长度一般为7-9个残基,但根据生物体和底物蛋白的不同,长度范围可以从4-25个残基不等。决定分解产物中肽段长度的机制,目前还没有完全弄清。
虽然具有催化活性的三个β亚基具有共同的降解机制,但它们对于底物的特异性却略有不同,分别为类胰凝乳蛋白酶型、类胰蛋白酶型和肽谷氨酰基肽水解型。这种对于底物特异性的差异是来自于靠近活性位点的局部残基与底物之间的相互作用的不同。每一个具有催化活性的β亚基也都含有一个降解所必需的保守的赖氨酸。
虽然蛋白酶体通常生成非常短的降解片断,但在一些情况下,这些降解产物自身是具有生物学活性的功能分子。特定的转录因子,包括哺乳动物的NF-κB(英语:NF-kB)复合物中的一个组分,合成后是以无活性的前体分子存在,在经过泛素化和蛋白酶降解后,才转变为活性分子。这种降解需要蛋白酶体剪切蛋白质的中间部分,而不是通常情况下的从蛋白质的一端开始的剪切。
有人提出,需要被剪切的中间部分为一个长的环(英语:loop(biochemistry)),位于蛋白表面,从而可以作为蛋白酶体的底物进入其内部孔道,而蛋白质的其他部分依然在孔道外,并不会被降解。在酵母蛋白中也发现了类似的现象;这种选择性降解被称为“受调控的泛素-蛋白酶体依赖的剪切”(regulated ubiquitin/proteasome dependent processing)。
非泛素依赖的降解
虽然大多数的蛋白酶体的底物必须在降解之前被泛素化,但仍然有一些例外的情况,尤其是在蛋白酶体参与蛋白质的翻译后处理过程中。一个主要的例子是蛋白酶体通过将p105(英语:p105)蛋白剪切为p50[英语:P50(biochemistry)]蛋白来激活NF-κB。一些由于存在无结构区域(英语:intrinsically unstructured proteins)而被推测具有不稳定性的蛋白质也可以通过非泛素依赖的途径被降解。鸟氨酸脱羧酶是最著名的非泛素依赖途径中蛋白酶体的底物。
对于关键的细胞周期调控因子,如P53蛋白的非泛素依赖的降解机制已经有报道,虽然p53蛋白也可以通过泛素依赖的途径被降解。此外,在一定的细胞应激条件下,结构不正常、错误折叠或者过度氧化的蛋白质也都会进入非泛素依赖的和非19S颗粒依赖的降解途径。
进化
大肠杆菌中的hslV(英语:HslV)(蓝色)和hslU(英语:HslU)复合物(红色)。这一由热休克蛋白组成的复合物被认为类似于现在蛋白酶体的祖先。
20S蛋白酶体在真核生物中广泛存在且必不可少。一些原核生物,包括许多古菌和细菌中的放线菌也含有20S蛋白酶体的同源体,即大多数细菌都含有的热休克基因hslV(英语:HslV)和hslU(英语:hslU),这两个基因所编码的蛋白质可以形成双层环状多聚体和ATP酶。一些研究者认为HslV蛋白很可能类似于20S蛋白酶体的祖先。一般来说,HslV蛋白对于细菌不是必要的,且并非所有的细菌都含有这一蛋白,而原生生物同时含有20S蛋白酶体和HslV蛋白系统。
序列分析显示,催化性的β亚基在进化过程中分化得比结构性的α亚基要早。表达20S蛋白酶体的细菌中,其β亚基与古菌以及真核生物的β亚基具有高度的序列相似性,而α亚基的序列相似程度则低得多。细菌中存在20S蛋白酶体可能是基因水平转移的结果,而真核生物中各亚基的分化则应是多次基因重复的结果。
作用
细胞周期控制
细胞周期进程是由一系列细胞周期蛋白依赖性激酶(CDK)来进行调控的,而CDK则是由细胞周期蛋白(cyclin)来激活。有丝分裂的细胞周期蛋白,在细胞中只有几分钟寿命,是所有已知的细胞内蛋白中寿命最短的。在CDK-cyclin复合物行使了它的功能之后,复合物中的cyclin就会被多泛素化并由蛋白酶体降解,从而保证了细胞周期的正常运转。尤其是在细胞退出有丝分裂期时,作为调控组分的周期蛋白B(英语:cyclin B)需要从有丝分裂促进因子上脱落下来,而这一解离过程依赖于蛋白酶体的参与。
细胞周期检控点,如G1期和S期之间的后限制点检查,也需要蛋白酶体降解周期蛋白A(英语:cyclin A),而cyclin A的泛素化由一个名为后期促进复合物(anaphase promoting complex,APC)的E3泛素连接酶来进行。APC蛋白和Skp1/Cul1/F-box蛋白复合物(即SCF复合物)是降解cyclin和控制检控点的两个关键调控因子;SCF复合物自身则由APC蛋白来调控,由于Skp2蛋白(SCF复合物中的转接蛋白)可以在G1期到S期的过渡期中抑制SCF复合物的活性,因此通过泛素化Skp2蛋白,APC蛋白就可以激活SCF复合物。
在植物中,生长素或植物激素的作用是调控植物生长的方向和向性,它们通过细胞信号通路来诱导一系列转录因子抑制蛋白(Aux/IAA蛋白)进入蛋白酶体降解途径。这些抑制蛋白由SCFTIR1蛋白或者由与auxin受体蛋白TIR1结合的SCF蛋白进行泛素化。Aux/IAA蛋白降解后,对auxin反应因子(ARF)家族的转录因子的抑制就被解除,从而诱导ARF基因的表达。
ARF被激活所导致的结果因植物类型和发育水平的不同而有所差异,但都参与了对根和叶脉生长的指导。ARF蛋白和Aux/IAA蛋白之间配对的特异性被认为是ARF的去抑制作用具有反应特异性的原因。
细胞凋亡
细胞内外的信号都能够诱导细胞凋亡或编程性细胞死亡。其结果是细胞内部的组分发生解构,这主要是由特定的蛋白酶半胱天冬酶来完成,但同时蛋白酶体也可能在细胞凋亡过程中扮演了多种重要角色。蛋白酶体参与细胞凋亡进程的推测是基于凋亡发生前,细胞中泛素化蛋白质以及E1、E2、E3在数量上的增加这一现象;并且,在细胞凋亡过程中,原本定位于细胞核的蛋白酶体被发现能够移位到调亡小泡的外膜。
蛋白酶体的抑制作用可以影响不同类型细胞的调亡诱导,在大多数已被研究的细胞类型中,抑制蛋白酶体可以促进细胞调亡。但一般而言,蛋白酶体并非是细胞调亡所必需的因子。而且,对于一些细胞系,特别是原代培养的静止和分化的细胞,如胸腺细胞和神经元细胞,暴露于蛋白酶体抑制剂反而阻止了细胞的凋亡。这一作用机制目前还不清楚,但有人推测这种现象只特异性地发生于静止状态的细胞或者这是由于促细胞凋亡激酶JNK(英语:JNK)的活性差异所导致的。由于蛋白酶抑制剂可以诱发处于快速分裂中的细胞(如癌细胞)的凋亡,因此一些蛋白酶抑制剂已经被开发并作为化疗药品被用于治疗癌症。
免疫系统
蛋白酶体直接参与了适应性免疫系统的运作,并在其中扮演着关键角色。肽类抗原是由主要组织相容性复合物(MHC)类型I蛋白传递到抗原呈递细胞表面。这些肽段是来自被蛋白酶体降解的侵入机体的病原体。虽然一般的蛋白酶体就可以参与这一进程,但实际上起主要作用的是一种特殊的复合物,其可以生成合适大小和成分的降解片断以供MHC结合。
这种复合物的组成蛋白的表达是由γ干扰素所诱导;当免疫反应发生时,这些蛋白质,包括11S调节颗粒(主要作用为调节MHC的结合肽段的产生)和特殊的β亚基(β1i、β2i、β5i,具有不同的底物特异性)的表达就会增加。这种由特殊的β亚基参与形成的复合物就被称为“免疫蛋白酶体”。另一种有所变化的β5亚基,β5t,在胸腺中表达,能够形成胸腺独有的“胸腺蛋白酶体”("thymoproteasome"),参与T细胞的发育调控。
MHC类型I蛋白的配基结合强度取决于配基C末端的组成,因为肽段配基是通过氢键和与MHC表面的"B pocket"近接触来结合的。许多MHC类型I蛋白趋向于结合疏水性残基,而免疫蛋白酶体复合物就可以更多地生成具有疏水性C末端的肽段。
由于蛋白酶体参与生成活性形势的NF-κB(英语:NF-kB)(一种抗凋亡和促炎症调控因子,调控细胞因子的表达),因此,蛋白酶体被认为与炎症反应和自身免疫性疾病相关。蛋白酶体活性水平的提高与包括红斑性狼疮和类风湿性关节炎在内的自身免疫性疾病相关。
蛋白酶体抑制剂
蛋白酶体抑制剂硼替佐米的化学结构。采用硼替佐米的化学疗法可以有效地治疗多发性骨髓瘤。
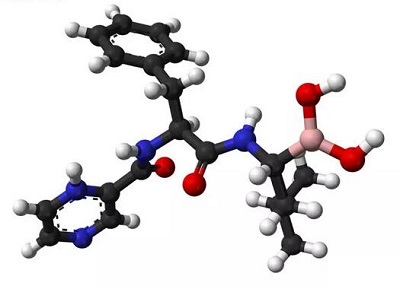 硼替佐米
硼替佐米硼替佐米结合于酵母蛋白酶体的核心颗粒上。硼替佐米分子位于图的正中;其中,粉色表示碳原子,蓝色表示氮原子,红色表示氧原子,黄色表示硼原子。环绕在硼替佐米分子周围的是蛋白质局部表面,其中蓝色的部分是发挥催化作用的苏氨酸残基,由于结合上硼替佐米,因而失去了催化活性。
蛋白酶体抑制剂对于人工培养的细胞具有有效的抗肿瘤活性,通过降解受调控的促生长细胞周期蛋白来诱导肿瘤细胞的凋亡。这种可以选择性地诱导肿瘤细胞凋亡的方法被证明在动物模型以及人体试验中都非常有效。硼替佐米是第一种用作化学治疗药物的蛋白酶体抑制剂,由千年制药公司(英语:Millennium Pharmaceuticals)开发,市场名称为Velcade。
硼替佐米主要被用于多发性骨髓瘤的治疗,值得一提的是,多发性骨髓瘤会导致血清中蛋白酶体水平的提高,而成功的化疗可以将蛋白酶体的水平恢复到正常范围。动物研究显示硼替佐米可能对死亡率极高的胰腺癌也有显着的临床效果。对于硼替佐米在治疗B细胞相关癌症的临床前和早期临床研究已经开始,特别是一些类型的非霍奇金氏淋巴瘤。
为了研究蛋白酶体抑制剂硼替佐米对急性白血病K562细胞株核因子κB(NF-κB)及细胞间黏附分子(ICAM-1)表达的影响,用含10%小牛血清的RPMI1640培养液体外培养K562细胞,用0、10、20、30、50、100nmol/L的硼替佐米干预K562细胞6小时后收集细胞。用SP免疫组织化学法测各组细胞NF-κB活性,并以反转录聚合酶链反应(RT-PCR)分析K562细胞ICAM-1表达的变化。
结果表明:各药物处理组K562细胞NF-κB的活性和ICAM-1的表达均明显受抑制,与对照组相比,存在显著性差异(p<0.05),且各组NF-κB活性与ICAM-1表达成正相关。结论:蛋白酶体抑制剂硼替佐米可能通过抑制NF-κB活性下调细胞间黏附分子ICAM-1的表达,这为白血病靶向治疗提供了一个新的途径。
利托那韦(英语:ritonavir),市场名称为Norvir,是用于治疗艾滋病的一种蛋白酶抑制剂。近期的研究发现,利托那韦不仅可以抑制蛋白酶,对蛋白酶体也有抑制作用,特别是对类胰凝乳蛋白酶型的蛋白酶体,但对类胰蛋白酶的蛋白酶体则有部分的促进作用。对于动物模型的研究表明利托那韦可能对神经胶质瘤细胞的生长有抑制作用。
将蛋白酶体抑制剂应用于自体免疫性疾病也大有前景。目前这一方面的研究主要是利用动物模型来进行。例如,在对植有人类皮肤的小鼠的研究中发现,用蛋白酶体抑制剂处理过后,原本因患牛皮癣而受损的皮肤所有减少;在一些啮齿动物模型中发现,蛋白酶体抑制剂对于哮喘也有一定的治疗作用。
蛋白酶体的标记和抑制作用也被用于在实验室中针对细胞中蛋白酶体活性所进行的“体外”(in vitro)和“体内”(in vivo)研究。实验室中最常用的抑制剂为乳胞素(英语:lactacystin),一种由链霉菌合成的天然产物。带有荧光基团的抑制剂也已经被开发应用于特异性标记组装好的蛋白酶体上的活性位点。
2004年诺贝尔化学奖的获奖主题就是蛋白质酶解在细胞中的重要性和泛素在酶解途径的作用,而三位获奖者为阿龙·切哈诺沃、阿夫拉姆·赫什科和欧文·罗斯。
参考资料
1.自噬与泛素化蛋白降解途径的分子机制及其功能·维普网
2.蛋白酶体抑制剂硼替佐米对K562细胞株NF-κB活性及ICAM-1表达的影响·维普网
")
")
")
")